Nematoda
Os Nematoda (nematódeos) são um filo de animais cilíndricos e alongados. Eram classificados, juntamente com outros grupos, no filo Nemathelminthes(nematelmintos), hoje obsoleto.
Possuem corpo não segmentado e revestido de cutícula resistente e quitinosa. Sistema digestivo completo, possuindo boca e ânus. O sistema nervoso é formado de um anel anterior, que circunda a faringe, e cordões nervosos longitudinais relacionados com aquele anel. O sistema locomotor é estruturado em camadas musculares longitudinais situadas logo abaixo da epiderme. As contrações desses músculos só permitem movimentos de flexão dorsoventral. Não há movimentos laterais. Todos os nematódeos são unissexuados (animais dióico), ou seja, têm sexos separados — machos e fêmeas distintos. Em alguns, há até nítido dimorfismo sexual (o macho é bem diferente da fêmea).
Não há estruturas flageladas nem ciliadas nesses animais. Nem mesmo os espermatozóides possuem flagelos. Eles se locomovem por meio de pseudópodos, com movimentos amebóides.
Os nematódeos não possuem sangue, sistema circulatório nem sistema respiratório. A respiração é anaeróbia. São animais triblásticos, protostômios, pseudocelomados. Seu corpo cilíndrico, alongado e não segmentado exibe simetria bilateral. Possuem sistema digestivo completo, sistemas circulatório e respiratório ausentes; sistema excretor composto por dois canais longitudinais (renetes-formato de H); sistema nervoso parcialmente centralizado, com anel nervoso ao redor da faringe.
Ecologicamente são muito bem-sucedidos, sendo tal fato demonstrado pela alta diversidade de espécies. Encontram-se em todos os habitats, terrestres, marinhos e de água doce e chegam a ser mais numerosos que os outros animais, tanto em número de espécies, como de indivíduos. Algumas espécies são microscópicas, enquanto a fêmea de uma espécie, parasita do cachalote chamada "Placentonema gigantissima" pode atingir 13 metros de comprimento.
Recentemente, aclamou-se que os nematódeos são uma das três principais radiações de organismos multicelulares que têm produzido a maioria das espécies do mundo, sendo as outras radiações os insetos e os fungos (Gaston, 1991).
| Nematoda | |||||||
|---|---|---|---|---|---|---|---|
 | |||||||
| Classificação científica | |||||||
| |||||||
| Classes | |||||||
| Adenophorea
Subclasse Enoplia
Subclasse Chromadoria Secernentea Subclasse Rhabditia Subclasse Spiruria Subclasse Diplogasteria | |||||||
Taxonomia e sistemáticaLineu (1758) descreveu algumas espécies de nematódeos, contidos na classe Vermes. Em 1808, Karl Rudolphi,[1] agrupou os atuais nematódeos e nematomorfosem Nematoidea, do grego antigo νῆμα (nêma, nêmatos, 'filamento') e -eiδἠς (-eidēs, 'espécie'). Os Nematoidea, junto com Acanthocephala, Trematoda e Cestoidea, formavam o grupo Entozoa.[2]
Nematoidea (ainda contendo nematomorfos) seria reclassificado como famíliaNematodes por Burmeister em 1837,[1] como ordem Nematoda por K. M. Diesing em 1861,[1] e como filo Nematoidea por Ray Lankester (1877).
A primeira diferenciação dos nematódeos dos nematomorfos, embora errônea, se deve a von Siebold (1843), com as ordens Nematoidea e Gordiacei (esta contendo alguns nematódeos). Vejdovsky (1866) separou adequadamente aos gordiáceos dos nematódeos, criando para os gordiáceos a classe Nematomorpha sob o filo Nemathelmintes.
Gegenbaur (1859) propôs o grupo Nemathelminthes, que incluía os Nematoidea, além de outros organismos. Grobben (1910) propôs o grupo Aschelminthes, com uma composição um pouco diferente de Nemathelminthes, incluindo Nematoda e Nematomorpha, dentre outros. Hyman (1951) reagrupou sob Pseudoceolomata os grupos Gastrotricha, Priapulida, Kinorhyncha, Nematoda, Nematomorpha e Rotifera. Tanto Nemathelminthes, Aschelminthes e Pseudoceolomata são hoje obsoletos.
Anatomia dos nematódeosOs nematódeos de vida livre são pequenos, geralmente menores do que 2,5 mm de comprimento e têm o corpo construído no mesmo plano fundamental, um cilindro quase perfeito, nu, delgado e alongado, com aspecto filiforme, em sua maioria, ou fusiforme. O animal é essencialmente um tubo dentro de outro tubo: o tubo externo é a parede corpórea, constituída, externamente, por uma cutícula complexa e, internamente, por uma camada de músculos longitudinais. O tubo interno é o trato digestivo, que é terminal na extremidade anterior mas subterminal posteriormente. Entre a parede e o tubo digestivo há a cavidade corpórea ou pseudoceloma, preenchida por líquido, que funciona como um "esqueleto hidrostático", além de favorecer a distribuição de nutrientes e recolher excretas, e na qual se encontram os órgãos reprodutores.
A epiderme é sincicial, ou seja, formada por uma massa celular multinucleada, ou celular, dependendo da espécie, e produz uma cutícula depositada externamente a ela. A cutícula é acelular, lisa, resistente e oferece proteção para o animal; em algumas formas, ela apresenta projecções que ajudam na locomoção. A cutícula tem de ser mudada para o animal crescer, um processo denominado ecdise - que coloca este filo no grupo dos Ecdysozoa, juntamente com os artrópodese outros filos.
Seus músculos são exclusivamente longitudinais, dispostos no sentido do comprimento do corpo. Isso faz com que a sua capacidade de locomoção seja mais limitada que a dos platelmintos. Os músculos são activados pelas cadeias nervosas, que se encontram ao longo de todo o corpo do animal, uma na região ventral e outra na dorsal. Ao contrário de outros animais, em que os nervos se ramificam para os músculos, nos nematódeos são os músculos que se ramificam para atingirem os cordões nervosos. Estes cordões ligam-se a um anel à volta da faringe e possuem vários gânglios adicionais perto da extremidade anterior, mas sem formar um verdadeiro cérebro. Nessa região encontram-se órgãos sensoriais reduzidos.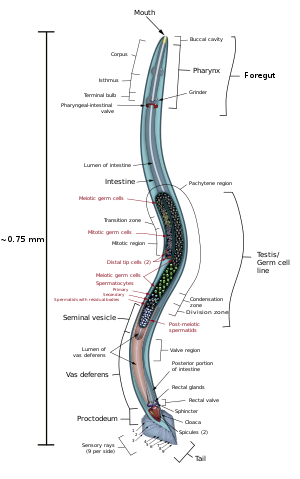

Sistema digestivo
O tubo digestivo dos nematódeos é completo, ou seja, possui um orifício de entrada de alimentos (a boca) e um outro orifício de saída de dejetos (o ânus) - são enterozoários completos.
Na boca, podem ser encontradas placas cortantes semelhantes a dentes, com as quais os nematódeos podem perfurar os tecidos de outros seres vivos. A faringe é musculosa e serve para esmagar os alimentos e também para os dirigir para o intestino, que não possui qualquer musculatura. O alimento é completamente digerido pelas enzimas que atuam sobre ele no interior do tubo digestivo, e os nutrientes são passados para a cavidade do corpo para serem distribuídos pelas células.
Muitos nematódeos de vida livre são carnívoros e se alimentam de pequenos animais ou de corpos de animais mortos. Os parasitas intestinais recebem o alimento já parcialmente digerido pelo hospedeiro
Sistema circulatório
Assim como os platelmintos, os nematódeos são avasculares (não possuem sistema circulatório). A cavidade corporal (o pseudoceloma) contém um líquido, e a contínua movimentação desse líquido, propiciada pela contração da musculatura longitudinal do corpo, permite uma relativa distribuição de materiais entre algumas partes do corpo.
Trocas gasosas
Eles não possuem órgãos respiratórios. As trocas gasosas acontecem na superfície corporal, por difusão. Os nematódeos de vida livre são aeróbicos e obtêm o oxigênio no meio onde vivem. Os parasitas são geralmente anaeróbicos e fazem fermentação. Dessa forma não requerem oxigênio e a maioria não elimina CO2, porque realizam a fermentação láctica, que não libera esse gás.
Sistema excretor
Os resíduos metabólicos são excretados a partir do líquido que ocupa o pseudoceloma, por meio de dois tubos longitudinais ligados por um menor, transversal. A distribuição desses tubos, no corpo, dá a eles o nome de tubos em "H". Os dois ramos longitudinais do sistema se abrem em orifícios próximos da boca.
Sistema nervoso
Possuem dois cordões nervosos que percorrem o corpo do animal, ventral ou longitudinalmente. Da faringe partem os cordões nervosos. O cordão nervoso dorsal é responsável pela função motora, enquanto a ventral é sensorial e motora, sendo considerada a mais importante.
Reprodução e crescimento
A maioria das espécies são dioicas, (realizam fecundação interna), ocorrendo em algumas nítido dimorfismo sexual: normalmente os machos são menores que as fêmeas, apresentam espinhos copulatórios e possuem a cauda encurvada.
Na cópula, os machos depositam os seus espermatozoides no poro genital das fêmeas. Os machos não possuem poro genital, e a saída dos espermatozoides ocorre pela cloaca.
Também são características exclusivas dos nematódeos a ausência de células ciliadas e os espermatozóides ameboides, sem flagelo, deslocando-se por pseudópodos.
A fecundação acontece dentro do corpo da fêmea (fecundação interna). Depois de fecundado, o zigoto se desenvolve dentro de um ovo com a casca resistente. Muitas espécies eliminam os ovos fecundados para o ambiente, onde as primeiras divisões se processam e o ovo se torna embrionado.
O ciclo evolutivo pode ser direto ou indireto, dependendo da formação de larvas por dentro ou fora dos ovos.





